Молекула РНК также полимер, мономерами которого является рибонуклеотиды, РНК представляет собой одноцепочную молекулу. Она построена таким же образом, как и одна из цепей ДНК. Нуклеотиды РНК похожи на нуклеотиды ДНК, хотя и не тождественны им. Их тоже четыре, и они состоят из осатков азотистого основания, пентозы и фосфорной кислоты. Три азотистых основания совершенно такие же, как в ДНК: А , Г и Ц . Однако вместо Т у ДНК в РНК присутствует близкое по строение пиримидиновое основание – урацил (У ). основное различие между ДНК и РНК – это характер углевода: в нуклотидах ДНК моносахарид – дезоксирибоза, а в РНК – рибоза. Связь между нуклеотидами осуществляется, как и в ДНК, через сахар и остаток фосфорной кислоты. В отличие от ДНК, содержание которой в клетках определенных организмов постоянно, содержание РНК в них колеблется. Оно заметно выше там, где происходит интенсивный синтез.
В отношении выполняемых функций различают несколько видов РНК.
Транспортная РНК (тРНК). Молекулы тРНк самые короткие: они состоят всего из 80-100 нуклеотидов. Молекулярная масса таких частиц равна 25-30 тыс. Транспортные РНК в основном содержатся в цитоплазме клетки. Функция их состоит в переносе аминокислот в рибосомы, к месту синтеза белка. Из общего содержания РНК клетк на долю тРНК приходится около 10%.
Рибосомная РНК (рРНК). Это крупные молекулы: в их состав входит 3-5 тыс. нуклеотидов, соотвественно их молекулярная масса достигает 1-1,5 млн. Рибосомные РНК составляют существенную часть рибосомы. Из общего содержания РНК в клетке на долю рРНК приходится около 90%.
Информационная РНК (иРНК), или матричная РНК (мРНК), содержится в ядре и цитоплазме. Функция ее состоит в переносе информации о структуре белка от ДНК к месту синтеза белка в рибосомах. На долю иРНК приходится примерно 0,5-1% от общего содержания РНК клетки. Размер иРНК колеблется в широких пределах – от 100 до 10000 нуклеотидов.
Все виды РНК синтезируются на ДНК, которая служитсвоего рода матрицей.
ДНК – носитель наследсвенной нформации.
Каждый белок представлен одной или несколькими полипиптидными цепями. Участок ДНК, несущий информацию об одной полипиптидной цепи, называют геном . Совокупность молекул ДНК клетки выполняет функцию носителя генетической информации. Генетическая информация передается как от материнской клетки дочерним клеткам, так и от родителей детям. Ген является единицей генетической , или наследственной, информации.
ДНК – носитель генетической информаци в клетке – непосредственного участия в синтезе белков не принимает. В клетках эукариот молекулы ДНК содержатся в хромосомах ядра и отделены ядерной оболочкой от цитоплазмы, где происходит синтез белков. К рибосомам – местам сборки белков – высылается из ядра несущий информацию посредник, способный пройти через поры ядерной оболочки. Таким посредником является информационная РНК (иРНК). По принципу комплементарности она синтезируется на ДНК при участие фермента, называемого РНК-полимеразой .

Информационная РНК – это однонитевая молекула, и транскрипция идет с одной цепи двунитевой молекулы ДНК. Она является копией не всей молекулы ДНК, а только части ее – одного гена у эукариот или группы рядом расположенных генов, несущих информацию о структуре белков, необходимых для выполнения одной функции, у прокариот. Такую группу генов называют опероном . В начале каждого оперона находится своего рода посадочная площадка для РНК-полимеразы, называемая промотором .это специфическая последовательность нуклеотидов ДНК, которую фермент «узнает» благодаря химическому сродству. Только присоединившись к промотору, РНК-полиммераза способна начать интез РНК. Доядя до конца оперона, фермент встречает сигнал (в виде определенной последоватльности нуклеотидов), означающий конец считывания. Готовая иРНК отходит от ДНК и направляется к месту синтеза белков.
В процессе транскрипции можно выделить четыре стадии: 1) связывание РНК -полимеразы с промотором; 2) инициация – начало синтеза. Оназаключается в образовании первой фосфодиэфирной связи между АТФ или ГТФ и вторым нуклеотидом синтезирующейся молекулы РНК; 3) элонгация – рост цепи РНК; т.е. последовательное присоединение нуклеотидов друг к другу в том порядке, в котором стоят комплементарные им нуклеотиды в транскрибируемой нити ДНК. Скорость элонгации 50 нуклеотидов в секунду; 4) терминация – завершение синтеза РНК.
Пройдя через поры ядерной оболочки, иРНК направляется к рибосомам, где осуществляется расшифровка генетической информации – перевод ее с «языка» нуклеотидов на «язык» аминокислот. Синтез полипептидных цепей по матрице иРНК, происходящий в рибосомах, называют трансляцией (лат. translation – перевод).
Аминокислоты, из котрых синтезируютсябелки, доставляются к рибосомам с помощью специальных РНК, называемых транспортными (тРНК). В клетке имеется столько же разных тРНК, сколько кодонов, шифрующих аминокислоты. На вершине «листа» каждой тРНК имеется последовательность трех нуклеотидов, комплементарных нуклеотидам кодона в иРНК. Ее называют антикодоном. Специальный фермент – кодаза – опознает тРНК и присоединяет к «черешку листа» аминокислоту – только ту, которая кодируется триплетом, комплементарным антикодону. На образование ковалентной связи между тРНК и «своей» аминокислотой затрачивается энергия одной молекулы АТФ.
 Для
того чтобы аминокислота включилась в
полипептидную цепь, она должна оторваться
от тРНК. Это становится возможным, когда
тРНК поступает на рибососму и антикодон
узнает свой кодон в иРНК. В рибосоме
имеется два участка для связывания двух
молекул тРНК. В один из этих участков,
называемый акцепторным
,
поступает тРНК с аминокислотой и
присоединяется к своему кодону (I).
Эта аминокислота присоединяет к себе
(акцептирует) растущую цепь белка (II)?
Между ними образуется пептидная связь.
тРНК, к которой теперь присоединяется
вместе с кодоном иРНК в донорный
участок рибосомы. В освободившийся
акцепторный участок приходит новая
тРНК, связанная с аминокислотой, которая
шифруется очередным кодоном (III).
Из донорного участка сюда вновь
переносится оторвавшаяся полипептидная
цепь и удлинняется еще на одно звено.
Аминокислоты в растущей цепи соединены
в той последовательности, в которой
расположены шифрующие их кодоны в иРНК.
Для
того чтобы аминокислота включилась в
полипептидную цепь, она должна оторваться
от тРНК. Это становится возможным, когда
тРНК поступает на рибососму и антикодон
узнает свой кодон в иРНК. В рибосоме
имеется два участка для связывания двух
молекул тРНК. В один из этих участков,
называемый акцепторным
,
поступает тРНК с аминокислотой и
присоединяется к своему кодону (I).
Эта аминокислота присоединяет к себе
(акцептирует) растущую цепь белка (II)?
Между ними образуется пептидная связь.
тРНК, к которой теперь присоединяется
вместе с кодоном иРНК в донорный
участок рибосомы. В освободившийся
акцепторный участок приходит новая
тРНК, связанная с аминокислотой, которая
шифруется очередным кодоном (III).
Из донорного участка сюда вновь
переносится оторвавшаяся полипептидная
цепь и удлинняется еще на одно звено.
Аминокислоты в растущей цепи соединены
в той последовательности, в которой
расположены шифрующие их кодоны в иРНК.
Когда на рибосоме оказывается один из трех триплетов (УАА, УАГ, УГА ), являющиеся «знаками препинания» между генами, ни одна тРНК не может занять место в акцепторном участке. Дело в том, что не существует антикодонов, комплементарных последовательностям нуклеотидов «знаков препинания». Оторвавшейся цепи не к чему присоединиться в акцепторном участке, и она покидает рибосому. Синтез белка завершен.
У прокариот синтез белков начинается с того, что кодон АУГ , расположенный на первом месте в копии с каждого гена, занимае в рибосоме такую позицию, что с ним взаимодействует антикодон особой тРНК, оединенной с формилментионином . Эта измененная форма аминокислоты метионина сразу попадает в донорный участок и выполняет роль заглавной буквы во фразе – с нее в бактериальной клетке начинается синтез любой полипептидной цепи. Когда триплет АУГ стоит не на первом месте, а внутри копии с гена, он кодирует аминокислоту метионин. После завершения синтеза полипептидной цепи формилметионин отщепляется от нее и в готовом белке отсуствует.
 Для
увеличения производства белков иРНК
часто проходит одновременно не по одной,
а по нескольким рибосомам. Акую структуру,
объединенную одной молекулой иРНК,
называют полисомой
.
На каждой рибосоме вэтом похожем на
нитку бус конвейере синтезируются
одинаковые белки.
Для
увеличения производства белков иРНК
часто проходит одновременно не по одной,
а по нескольким рибосомам. Акую структуру,
объединенную одной молекулой иРНК,
называют полисомой
.
На каждой рибосоме вэтом похожем на
нитку бус конвейере синтезируются
одинаковые белки.
Аминокислоты бесперебойно поставляются к рибосомам с помощью тРНК. Отдав аминокислоту, тРНК покидает рибосому и с помощью кодазы соединяется. Высокая слаженность всех «служб комбината» по производсву белов позволяет в течении нескольких секунд синтезировать полипептидные цепи, состоящие из сотен аминокислот.
Свойства генетического кода. Благодаря процессу транскрипции в клетке осуществляется передача информации от ДНК к белку
ДНК → иРНК → белок
Генетическая информация, содержащаяся в ДНК и в иРНК, заключена в последовательности расположения нуклеотидов в молекулах.
 Каким
же образом происходит перевод информации
с «языка» нуклеотидов на «язык»
аминокислот? Такой перевод осуществляется
с помощью генетического кода. Код,
или шифр
,
- это система символов для перевода
одной формы информации в другую.
Генетический
код
–это
система записи информации о
последовательности расположения
аминокислот в белках с помощью
последовательности расположения
нуклеотидов в иРНК.
Каким
же образом происходит перевод информации
с «языка» нуклеотидов на «язык»
аминокислот? Такой перевод осуществляется
с помощью генетического кода. Код,
или шифр
,
- это система символов для перевода
одной формы информации в другую.
Генетический
код
–это
система записи информации о
последовательности расположения
аминокислот в белках с помощью
последовательности расположения
нуклеотидов в иРНК.
Какими же свойствами обладает генетический код?
Код триплетен . В состав РНК входят четыре нуклеотида: А, Г, Ц, У. Если бы мы пытались обозначить одну аминокислоту одним нуклеотидом, то 16 из 20 аминокислот остались бы не зашифрованы. Двух буквенный код позволил бы зашифровать 16 аминокислот. Природа создала трехбуквенный, или триплетный, код. Это означает, что каждая из 20 аминокислот зашифрована последовательностью из трех нуклеотидов, называемой триплетом или кодоном.
Код вырожден. Это означает, что каждая аминокислота шифруется более чем одним кодоном. Исключения: метеонин и триптофан, каждая из которых кодируется одним триплетом.
Код однозначен. Каждый кодон шифрует только одну аминокислоту.
Между генами имеется «знаки препинания». В печатном тексте в конце каждой фразы стоит точка. Несколько связанных по смыслу фраз составляют абзац. На языке генетической информации таким абзацем являетсяоперон и комплементарная ему иРНК. Каждый ген в опероне прокариот или отдельный ген эукариот кодирует одну полипептидную цепочку – фразу. Так как в ряде случаев по матрице иРНК последовательно создается несколько разных полипептидных цепей, они должны быть отделены друг от друга. Для этого в генетическом годе имеются три специальных триплета – УАА, УАГ, УГА, каждый из которых обозначает прекращение синтеза одной полипептидной цепи. Таким образом, эти триплеты выполняют функцию знаков препинания. Они находятся в конце каждого гена.
Внутри гена нет «знаков препинания».
Код универсален. Генетический код един для всех живущих на Земле существ. У бактерий и грибов, пшеницы и хлопка, рыб и червей, лягушек и человека одни и те же триплеты кодируют одни и те же аминокислоты.

Принципы репликации ДНК. Преемственность генетического материала в поколениях клеток и организмов обеспечивается процессом репликации – удвоения молекул ДНК. Этот сложный процесс осуществляется комплексом нескольких ферментов и не обладающих каталитической активностью белов, необходимых для придания полинуклеотидным цепям нужной конформации. В результате репликации образуются две идентичные двойные спирали ДНК. Эти так называемые дочерние молекулы ничем не отличаются друг от друга и от исходной материнской молекулы ДНК. Репликация происходит в клетке перед делением, поэтому каждая дочерняя клетка получает точно такие же молекулы ДНК, какие имела материнская клетка. Процесс репликации основан на ряде принципов:
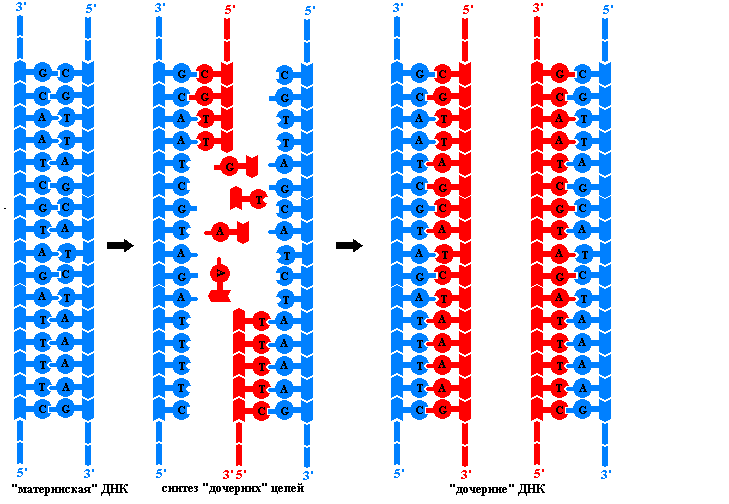
Только
в этом случае ДНК-полимеразы способна
двигаться по материнским нитям и
использовать их в качестве матриц для
безошибочного синтеза дочерних цепей.
Но полное раскручивание спиралей,
состоящих из многих миллионов пар
нуклеотидов, сопряжено со столь
значительным числом вращений и такими
энергетическими затратами, которые
невозможны в условиях клетки. Поэтому
репликация у эукариот начинается
одновременно в некоторых местах молекулы
ДНК. Участок между двумя точками, в
которых начинается синтез дочерних
цепей, называют репликоном
.
Он является единицей
репликации.


В каждой молекуле ДНК эукариотической клетки имеется много репликонов. В каждом репликоне можно видеть репликативную вилку – ту часть молекулы ДНК, которая под действием специальных ферментов уже расплелась. Каждая нить в вилке служит матрицей для синтеза комплементарной дочерней цепи. В ходе репликации вилка перемещается вдоль материнской молекулы, при этом расплетаются новые участки ДНК. Так как ДНК-полимеразы могут двигаться лишь в одном направлении вдоль матричных нитей, а нити ориентированы антипараллельно, то в каждой вилке одновременно ведут синтез два разных ферментативных комплекса. Причем в каждой вилке одна дочерняя (лидирующая) цепь растет непрерывно, а другая (отстающая) синтезируется отдельными фрагментами длинной в несколько нуклеотидов. Такие ферменты, названые в честь открывшего их японского ученого фрагментами Оказаки , сшиваются ДНК-лигазой, образуя непрерывную цепь. Механизм образования дочерних цепей ДНК фрагментами называют прерывистыми.
Потребность в затравке ДНК-полимераза не способна начинать синтез лидирующей цепи, ни синтез фрагментов Оказаки отстающей цепи. Она может лишь наращивать уже имеющуюся полинуклеотидную нить, последовательно присоединяя дезоксирибонуклеотиды к ее 3’-ОН концу. Откуда же берется начальный 5’-концевой участок растущей цепи ДНК? Его синтезирует на матрице ДНК особая РНК-полимераза, называемая праймазой (англ. Primer – затравка). Размер рибонуклеотидной затравки невелик (менее 20 нуклеотидов) в сравнении с размером цепи ДНК, образуемой ДНК-поимеразой. Выполнившая сво. Функци. РНК-затравка удаляется специальным ферментом, а образованная при эом брешь заделывается ДНК-полимеразой, использующей в качестве затравки 3’-ОН конец соседнего фрагмента Оказаки.
Проблема недорепликации концов линейных молекул ДНК. Удаление крайних РНК-праймеров, комплементрных 3’-концам обеих цепей линейной материнской молекулы ДНК, приводит к тому, что дочерние цепи оказываются короче 10-20 нуклеотидов. В этом и заключается проблема недорепликации концов линейных молекул.
Проблема недорепликации 3’-концов линейных молекул ДНК решается эукариотическими клетками с помощью специального фермента – теломеразы .
Теломераза является ДНК-полимеразой, достраивающей 3’-концылинейных молекул ДНК хромосом короткими повторяющимися последовательностями. Они, располагаясь друг за другом, образуют регулярную концевую структуру длинной до 10 тыс. нуклеотидов. Помимо белковой части, теломераза содержит РНК, выполняющую роль матрицы для наращивания ДНК повторами.
Схема удлинения концов молекул ДНК. Сначала происходит комплементарное связывание выступающего конца ДНК с матричным участком теломеразной РНК, затем теломераза наращивает ДНК, используя в качестве затравки ее 3’-ОН конец, а в качестве матрицы – РНК, входящую в состав фермента. Эта стадия называется элонгацией. После этого происходит транслокация, т.е. перемещение ДНК, удлиненной на один повтор, относительно фермента. Следом идет элонгация и очередная транслокация.
В результате образуются специализированные концевые структуры хромосом. Они состоят из многократно повторенных коротких последовательностей ДНК и специфических белков.
Транспортная РНК, тРНК -рибонуклеиновая кислота, функцией которой является транспортировка АК к месту синтеза белка. Имеет типичную длину от 73 до 93 нуклеотидов и размеры около 5 нм. тРНК также принимают непосредственное участие в наращивании полипептидной цепи, присоединяясь - будучи в комплексе с аминокислотой - к кодону мРНК и обеспечивая необходимую для образования новой пептидной связи конформацию комплекса. Для каждой аминокислоты существует своя тРНК. тРНК является одноцепочечной РНК, однако в функциональной форме имеет конформацию «клеверного листа». АК ковалентно присоединяется к 3"-концу молекулы с помощью специфичного для каждого типа тРНК фермента аминоацил-тРНК-синтетазы. На участке C находится антикодон, соответствующий АК-те. тРНК синтезируются обычной РНК-полимеразой в случае прокариот и РНК-полимеразой III в случае эукариот. Транскрипты генов тРНК подвергаются многостадийному процессингу, который приводит к формированию типичной для тРНК пространственной структуры.
Процессинг тРНК включает 5 ключевых этапов:
удаление 5"-лидерной нуклеотидной последовательности;
удаление 3"-концевой последовательности;
добавление последовательности CCA на 3"-конец;
вырезание интронов (у эукариот и архей);
модификации отдельных нуклеотидов.
Транспорт тРНК осущ-ся по Ran-зависимому пути при участии транспортного фактора экспортина t, кот.распознаёт характерную вторичную и третич.стр-ру зрелой тРНК: короткие двуспиральные участки и правильно процессированные 5"- и 3"-концы. Такой механизм обеспечивает экспорт из ядра только зрелых тРНК.
62. Трансляция – узнавание кодона мРНК
Трансляция – это осуществляемый рибосомами синтез белка из аминокислот на матрице мРНК (или и РНК). Cоставляющие элементы процесса трансляции: аминокислоты, тРНК, рибосомы, мРНК, ферменты для аминоацилирования тРНК, белковые факторы трансляции (белковые факторы инициации, элонгации, терминации - специфические внерибосомные белки, необходимые для процессов трансляции), источники энергии АТФ и ГТФ, ионы магния (стабилизируют структуру рибосом). В синтезе белка участвует 20 аминокислот. Чтобы аминокислота «узнала» свое место в будущей полипепетидной цепи, она должна связаться с транспортной РНК (тРНК), выполняющей адапторную функцию. Затем тРНК, связавшаяся с аминокислотой «узнает» соответствующий кодон на мРНК. Узнавание кодона мРНК:
Взаимодействие кодон - антикодон основано на принципах комплементарности и антипараллельности:
3’----Ц - Г- А*------5’ Антикодон тРНК
5’-----Г- Ц- У*------3’ Кодон мРНК
Гипотеза качания (wobble) была предложена Ф. Криком:
3′- основание кодона мРНК имеет нестрогое спаривание с 5′- основанием антикодона тРНК: например, У (мРНК) может взаимодействовать с А и Г (тРНК)
Некоторые тРНК могут спариваться с более, чем одним кодоном.
63. Характеристика составляющих элементов процесса трансляции. Трансляция (translatio-перевод)- процесс синтеза белка из аминокислот на матрице информационной (матричной) РНК (иРНК, мРНК), осуществляемый рибосомой.
Синтез белка является основой жизнедеятельности клетки. Для осуществления этого процесса в клетках всех организмов имеются спец.органеллы-рибосомы - рибонуклеопротеидные комплексы, построенные из 2 субъединиц: большой и малой. Функция рибосом заключается в узнавании трёхбуквенных (трехнуклеотидных) кодонов мРНК, сопоставлении им соответствующих антикодонов тРНК, несущих аминокислоты , и присоединении этих аминокислот к растущей белковой цепи. Двигаясь вдоль молекулы мРНК, рибосома синтезирует белок в соответствии с информацией, заложенной в молекуле мРНК.
Для узнавания АК-т в клетке имеются специальные «адаптеры», молекулы транспортной РНК (тРНК). Эти молекулы, имеющие форму клеверного листа, имеют участок (антикодон), комплементарный кодону мРНК, а также другой участок, к которому присоединяется аминокислота, соответствующая этому кодону. Присоединение аминокислот к тРНК осуществляется в энерго-зависимой реакции ферментами аминоацил-тРНК-синтетазами, а получившаяся молекула называется аминоацил-тРНК. Таким образом, специфичность трансляции определяется взаимодействием между кодоном мРНК и антикодоном тРНК, а также специфичностью аминоацил-тРНК-синтетаз, присоединяющих аминокислоты строго к соответствующим им тРНК (например, кодону GGU будет соответствовать тРНК, содержащая антикодон CCA, а к этой тРНК будет присоединяться только АК глицин).
Прокариотическая рибосома
5S и 23S рРНК 16S рРНК
34 белка 21 белок
Рибосомы прокариот имеют константу седиментации 70S, отчего получили название 70S-частиц. Они построены из двух неодинаковых субчастиц: 30S- и 50S-субъединиц. Каждая субъединица представляет комплекс рРНК и рибосомных белков.
30S-частица содержит одну молекулу 16S-рРНК и в большинстве случаев по одной молекуле белка из более 20 видов (21) . 50S-субъединица состоит из двух молекул рРНК (23S и 5S). В ее состав входят более 30 различных белков (34), также представленных, как правило, одной копией. Большая часть рибосомальных белков выполняет структурную функцию.
Эукариотическая рибосома
5S; 5,8S и 28S рРНК 18S рРНК
не менее 50 белков не менее 33 белков
Рибосома состоит из большой и малой субъединиц. Основу структуры каждой субъединицы составляет сложн образом свернутая рРНК. К каркасу из рРНК прикрепл рибосомн белки.
Коэффициент седиментации полной эукариотической рибосомы составляет около 80 единиц Сведберга (80S), а коэффициент седиментации ее субчастиц составляет 40S и 60S.
Меньшая 40S-субчастица состоит из одной молекулы 18S-рРНК и 30-40 белковых молекул. Большая 60S-субчастица содержит три типа рРНК с коэффициентами седиментации 5S, 5,8S и 28S и 40-50 белков (например, рибосомы гепатоцитов крысы включают 49 белков).
Функциональные участки рибосом
Р – пептидильный участок для пептидил тРНК
А – аминоацильный участок для аминоацил тРНК
Е – участок для выхода тРНК из рибосомы
Рибосома содержит 2 функциональных участка для взаимодействия с тРНК: аминоацильный (акцепторный) и пептидильный (донорный). Аминоацил-тРНК попадает в акцепторный участок рибосомы и взаимодействует с образованием водородных связей между триплетами кодона и антикодона. После образования водородных связей система продвигается на 1 кодон и оказывается в донорном участке. Одновременно в освободившемся акцепторном участке оказывается новый кодон, и к нему присоединяется соответствующий аминоацил-т-РНК.
Рибосомы: строение, финкция
Рибосомы являются цитоплазматическими центрами биосинтеза белка. Они состоят из большой и малой субъединиц, различающихся коэффициентами седиментации (скоростью осаждения при центрифугировании), выражаемые в единицах Сведберга – S.
Рибосомы присутствуют в клетках как эукариот, так и прокариот, поскольку выполняют важную функцию в биосинтезе белков. В каждой клетке имеются десятки, сотни тысяч (до нескольких миллионов) этих мелких округлых органоидов. Это округлая рибонуклеопротеиновая частица. Диаметр ее составляет 20-30 нм. Состоит рибосома из большой и малой субъединиц, различающихся коэффициентами седиментации (скоростью осаждения при центрифугировании), выражаемые в единицах Сведберга – S. Эти субъединицы объединяются в присутствии нити м-РНК (матричной, или информационной, РНК). Комплекс из группы рибосом, объединенных одной молекулой м-РНК наподобие нитки бус, называется полисомой . Эти структуры либо свободно расположены в цитоплазме, либо прикреплены к мембранам гранулярной ЭПС (в обоих случаях на них активно протекает синтез белка).
Полисомы гранулярной ЭПС образуют белки, выводимые из клетки и используемые для нужд всего организма (например, пищеварительные ферменты, белки женского грудного молока). Кроме этого, рибосомы присутствуют на внутренней поверхности мембран митохондрий, где также принимают активное участие в синтезе белковых молекул.
В цитоплазме клеток содержатся три основных функциональных вида РНК:
- матричные РНК (мРНК), выполняющие функции матриц белкового синтеза;
- рибосомные РНК (рРНК), выполняющие роль структурных компонентов рибосом;
- транспортные РНК (тРНК), участвующие в трансляции (переводе) информации мРНК в последовательность аминокислот молекулы белка.
В ядре клеток обнаруживают ядерную РНК, составляющую от 4 до 10% от суммарной клеточной РНК. Основная масса ядерной РНК представлена высокомолекулярными предшественниками рибосомных и транспортных РНК. Предшественники высокомолекулярных рРНК (28 S, 18 S и 5 S РНК) в основном локализуются в ядрышке.
РНК является основным генетическим материалом у некоторых вирусов животных и растений (геномные РНК). Для большинства РНК вирусов характерна обратная транскрипция их РНК генома, направляемая обратной транскриптазой.
Все рибонуклеиновые кислоты представляют собой полимеры рибонуклеотидов, соединенных, как в молекуле ДНК, 3",5"-фосфорнодиэфирными связями. В отличие от ДНК, имеющей двухцепочечную структуру, РНК представляет собой одноцепочечные линейные полимерные молекулы.
Строение мРНК. мРНК - наиболее гетерогенный в отношении размеров и стабильности класс РНК. Содержание мРНК в клетках составляет 2-6% от общего количества РНК. мРНК состоят из участков - цистронов, определяющих последовательность аминокислот в кодируемых ими белках.
Строение тРНК . Транспортные РНК выполняют функции посредников (адаптеров) в ходе трансляции мРНК. На их долю приходится примерно 15% суммарной клеточной РНК. Каждой из 20 протеиногенных аминокислот соответствует своя тРНК. Для некоторых аминокислот, кодируемых двумя и более кодонами, существуют несколько тРНК. тРНК представляют собой сравнительно небольшие одноцепочечные молекулы, состоящие из 70-93 нуклеотидов. Их молекулярная масса составляет (2,4-3,1) .104 кДа.
Вторичная структура тРНК формируется за счет образования максимального числа водородных связей между внутримолекулярными комплементарными парами азотистых оснований. В результате образования этих связей полинуклеотидная цепь тРНК закручивается с образованием спирализованных ветвей, заканчивающихся петлями из неспаренных нуклеотидов. Пространственное изображение вторичных структур всех тРНК имеет форму клеверного листа.
В «клеверном листе» различают четыре обязательные ветви , более длинные тРНК, кроме того, содержат короткую пятую (дополнительную) ветвь . Адапторную функцию тРНК обеспечивают акцепторная ветвь, к 3"-концу которой присоединяется эфирной связью аминокислотный остаток, и противостоящая акцепторной ветви антикодоновая ветвь, на вершине которой находится петля, содержащая антикодон. Антикодон представляет собой специфический триплет нуклеотидов, который комплементарен в антипараллельном направлении кодону мРНК, кодирующему соответствующую аминокислоту.
Т-Ветвь, несущая петлю псевдоуридина (ТyС-петлю), обеспечивает взаимодействие тРНК с рибосомами.
Д-ветвь, несущая дегидроуридиновую петлю, обеспечивает взаимодействие тРНК с соответствующей аминоацил-тРНК-синтетазой.
Вторичная структура тРНК
Функции пятой дополнительной ветви пока мало исследованы, вероятнее всего она уравнивает длину разных молекул тРНК.
Третичная структура тРНК очень компактна и образуется путем сближения отдельных ветвей клеверного листа за счет дополнительных водородных связей с образованием L-образной структуры «локтевого сгиба» . При этом акцепторное плечо, связывающее аминокислоту, оказывается расположенным на одном конце молекулы, а антикодон - на другом.

Третичная структура тРНК (по А.С. Спирину)
Строение рРНК и рибосом . Рибосомные РНК формируют основу, с которой связываются специфические белки при образовании рибосом. Рибосомы - это нуклеопротеиновые органеллы, обеспечивающие синтез белка на мРНК. Число рибосом в клетке очень велико: от 104 у прокариот до 106 у эукариот. Локализуются рибосомы главным образом в цитоплазме, у эукариот, кроме того, в ядрышке, в матриксе митохондрий и строме хлоропластов. Рибосомы состоят из двух субчастиц: большой и малой. По размерам и молекулярной массе все изученные рибосомы делят на 3 группы - 70S рибосомы прокариот (S-коэффициент седиментации), состоящие из малой 30S и большой 50S субчастиц; 80S рибосомы эукариот, состоящие из 40S малой и 60S большой субчастиц.
Малая субчастица 80S рибосом образована одной молекулой рРНК (18S) и 33 молекулами различных белков. Большая субчастица образована тремя молекулами рРНК (5S, 5,8S и 28S) и примерно 50 белками.
Вторичная структура рРНК образуется за счет коротких двуспиральных участков молекулы - шпилек (около 2/3 рРНК), 1/3 - представлена однотяжевыми участками , богатыми пуриновыми нуклеотидами.
Все тРНК имеют общие черты как в их первичной структуре, так и в способе складывания полинуклеотидной цепи во вторичную структуру за счет взаимодействий между основаниями нуклеотидных остатков.
Первичная структура тРНК
тРНК - относительно небольшие молекулы, длина их цепей варьирует от 74 до 95 нуклеотидных остатков. Все тРНК имеют одинаковый 3"-конец, построенный из двух остатков цитозина и одного - аденозина (CCA-конец). Именно 3"-концевой аденозин связывается с аминокислотным остатком при образовании аминоацил-тРНК. CCA-конец присоединяется ко многим тРНК с помощью специального фермента. Нуклеотидный триплет, комплементарный кодону для аминокислоты (антикодон), находится приблизительно в середине цепи тРНК. В отдельных положениях последовательности практически у всех видов тРНК встречаются одни и те же (консервативные) нуклеотидные остатки. В некоторых положениях могут находиться или только пуриновые, или только пиримидиновые основания (их называют полуконсервативными остатками).
Для всех молекул тРНК характерно присутствие большого числа (до 25% всех остатков) разнообразных модифицированных нуклеозидов, часто называемых минорными. Они образуются в различных местах молекул, во многих случаях четко определенных, в результате модификации обычных нуклеозидных остатков с помощью специальных ферментов.
Вторичная структура тРНК
складывания цепи во вторичную структуру происходит за счет взаимокомплементарности участков цепи. Три фрагмента цепи оказываются комплементарными при складывании их на себя, образуя шпилькообразные структуры. Кроме того, 5"-конец комплементарен участку, близкому к 3"-концу цепи, при их антипараллельном расположении; они формируют так называемый акцепторный стебель. В результате образуется структура, характеризующаяся наличием четырех стеблей и трех петель, которая получила название "клеверного листа". Стебель с петлей формируют ветвь. Внизу расположена антикодоновая ветвь, содержащая антикодоновый триплет в составе своей петли. Слева и справа от нее расположены D- и T-ветви, соответственно названные так из-за присутствия в их петлях необычных консервативных нуклеозидов дигидроуридина (D) и тимидина (T). Нуклеотидные последовательности всех изученных тРНК могут быть сложены в аналогичные структуры. В дополнение к трем петлям клеверного листа в структуре тРНК выделяют также дополнительную, или вариабельную, петлю (V-петлю). Ее размеры резко различаются у разных тРНК, варьируя, от 4 до 21 нуклеотида, а по последним данным, и до 24 нуклеотидов.
Пространственная (третичная) структура тРНК
За счет взаимодействия элементов вторичной структуры формируется третичная структура, которая получила название L-формы из-за сходства с латинской буквой L (рис. 2 и 3). За счет стэкинга оснований акцепторный стебель и T-стебель клеверного листа образуют одну непрерывную двойную спираль, а два других стебля - антикодоновый и D - другую непрерывную двойную спираль. При этом D- и T-петли оказываются сближенными и скрепляются между собой путем образования дополнительных, часто необычных пар оснований. В образовании этих пар, как правило, принимают участие консервативные или полуконсервативные остатки. Аналогичные третичные взаимодействия скрепляют и некоторые другие участки L-структуры
Молекулы РНК в отличие от ДНК построены из одной полинуклеотидной цепи. Однако в этой цепи (для рРНК и мРНК) имеются комплементарные друг другу участки, которые могут взаимодействовать, образуя двойные спирали. При этом соединяются водородными связями нуклеотидные пары А-У и Г-Ц. Такие спирализованные участки (их называют шпильками) обычно содержат небольшое количество нуклеотидных пар (до 20-30) и чередуются с неспирализованными участками.
Характерную вторичную структуру имеют тРНК. Они содержат четыре спирализованных участка и три (четыре) одноцепочные петли. При изображении такой структуры на плоскости получается фигура, называемая «клеверным листом» (рис. справа).
Рис.. Вторичная (справа) и третичная (слева) структура тРНК
Все несколько десятков разных тРНК клетки имеют общий план пространственной структуры, но различаются в деталях. В тРНК выделяют следующие структурные участки.
1. Акцепторный конец - во всех типах тРНК имеет состав ЦЦА. К гидроксилу З"-ОН аденозина карбоксильной группой присоединяется аминокислота, которую данная тРНК доставляет к рибосомам, где происходит синтез белка.
2. Антикодоновая петля - содержит специфический для каждой тРНК триплет нуклеотидов (антикодоны). Антикодон комплементарен кодону мРНК. Кодон-антикодоновое взаимодействие определяет порядок чередования аминокислот в белковой молекуле при синтезе ее на рибосомах.
3. Псевдоуридиловая петля (Г,С) - участвует в связывании тРНК с рибосомой.
4. Дигидроуридиловая (D) петля необходима для связывания с ферментом аминоацил-тРНК-синтетзой, которая участвует в узнавании аминокислотой своей тРНК.
5. Добавочная петля - разная у разных тРНК.
Третичная структура рнк и днк
Пространственная конфигурация спирализованной полинуклеотидной цепи (третичная структура) достаточно полно выяснена для молекул РНК. Установлено, что нативные молекулы тРНК имеют примерно одинаковую третичную структуру, которая отличается от плоской структуры «клеверного листа» (вторичная структура) большей компактностью за счет складывания различных частей молекулы (см. рис выше).
Для рРНК и мРНК возможно существование, а зависимости от концентрации солей и температуры, трех видов третичной структуры (рис. ниже). Первый - рыхлый беспорядочный клубок или распрямленная цепь (при повышении температуры и отсутствии солей). Второй вариант - компактный клубок с двуспиральными участками (высокая ионная сила, комнатная температура). Третий вид - компактная палочка с упорядочение ориентированными двуспиральными участками (низкая ионная сила, комнатная температура). Все три типа третичной структуры РНК связаны взаимными переходами.
Третичная структура ДНК зависит от того, сколько цепочек полинуклеотидов (одна или две) в ДНК. В ряде вирусов обнаружены одноцепочечные ДНК линейной и кольцевой формы. Двуцепочечные спиралевидные молекулы ДНК также могут существовать в линейной и кольцевой форме; образование последней вызвано ковалентным соединением их открытых концов.

Рис. Третичная структура: А - ДНК: 1 - линейная одноцепочечная бактериофаг ФХ174 (и др. вирусов); 2 - кольцевая одноцепочечная ДНК вирусов и митохондрий; 3 - кольцевая двойная спираль ДНК; Б - РНК: 1 - рыхлый клубок или распрямленная цепь; 2 - компактная палочка; 3 - компактный клубок
Кроме того, полагают, что биспиральные молекулы ДНК существуют в хромосомах в виде вторично спирализованных фрагментов, связанных друг с другом (суперспираль). Поэтому молекулярный вес нативной ДНК достигает нескольких сотен миллионов. Следовательно, молекулы с молекулярной массой 10.000.000 являются субъединицами более крупных молекулярных образований (третичная структура). Именно суперспирализация обеспечивает экономную упаковку огромной молекулы ДНК в хромосоме: вместо 8 см длины, которую она могла бы иметь в вытянутой форме, она занимает всего 5 нм.


